CASE STUDY IN MOLECULAR EVOLUTION NO. 5
Written by Harold B. White 9/93, Revised most recently Oct. 2000
C-647 BIOCHEMICAL EVOLUTION, FALL 2000
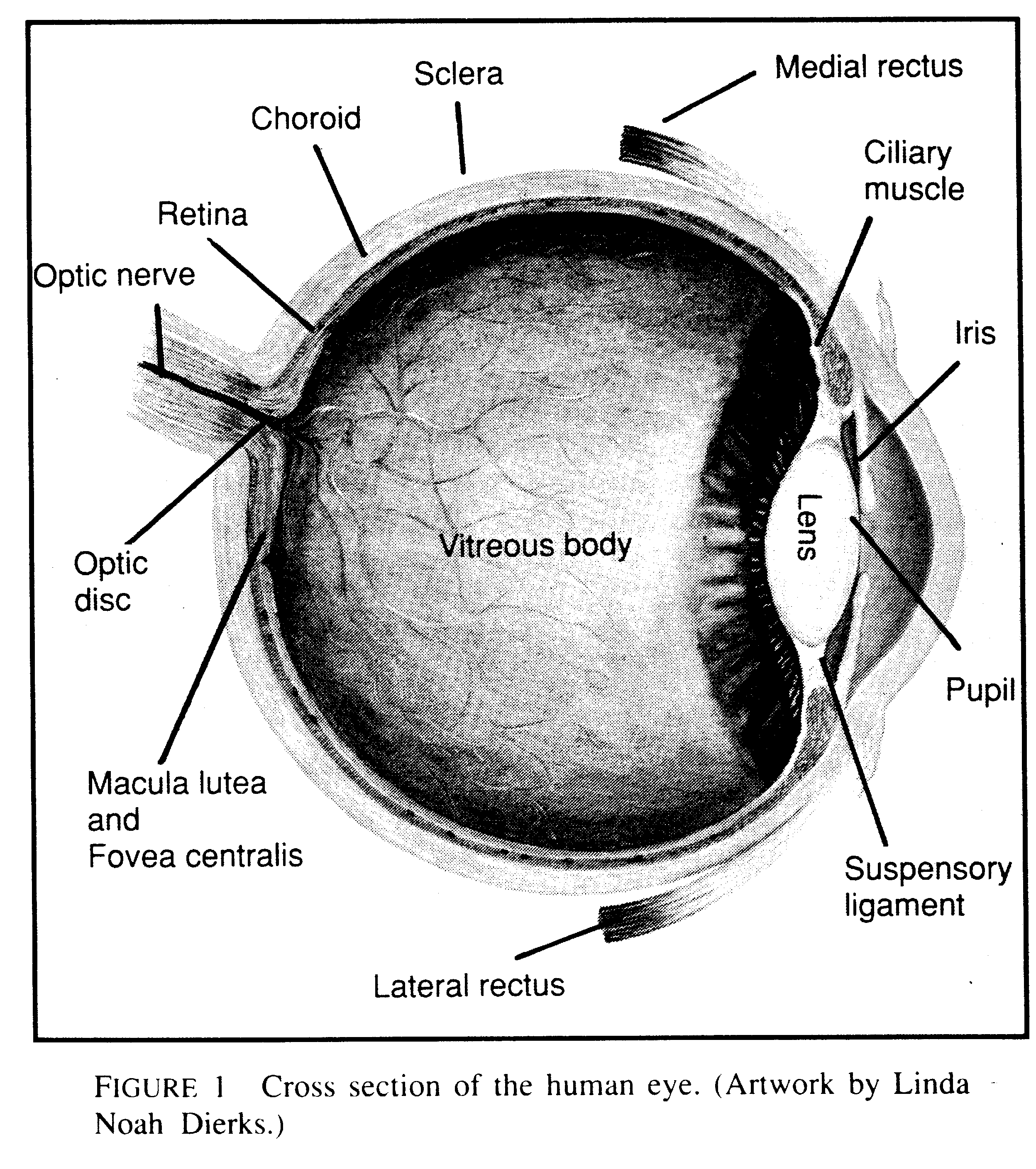
ATo suppose that the eye with all its inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for correction of spherical and chromatic aberration could have been formed by natural selection, seems, I freely confess, absurd in the highest degree.@ Charles Darwin (1)
Perhaps because of Darwin=s admission quoted above, the evolution of eyes has had unusual fascination for both supporters and critics of evolutionary theory. Most of this discussion has related to the macroscopic elements of eyes. Recently, with the ability to analyze proteins and genes in molecular detail, the evolution of the eye has assumed dimensions never Aenvisioned@ by Darwin. The following begins an interesting biochemical story in the evolution of eyes.
Why should any organism or cell need more than one enzyme to catalyze a particular reaction? In the 1960's "isozymes" were hot in biochemical research. Equipped with starch gel electrophoresis and enzyme-specific staining methods, biochemists and geneticists discovered that most intracellular enzymes are composed of multiple, but not always identical, subunits. They showed that homologous genes and their alleles encode the non-identical interchangeable subunits. Isozymes and allozymes showed up everywhere. Articles describing subunit structures, electrophoretic variants, and allele frequencies filled the journals. From these studies, it became apparent that isozymes arise from gene duplication and expression of the resulting independently evolving homologous genes. Most researchers assumed that duplicated genes were retained by selection for subtle, although rarely identified, functional differences in the encoded isozymes. Lactate dehydrogenase, perhaps the archetypical isozyme system of that era, conformed nicely to the model (2, 3).
L-Lactate dehydrogenase (LDH), an important glycolytic enzyme, is a tetrameric protein composed of identical or structurally similar subunits (4). In vertebrates, three homologous genes on different chromosomes encode 35 kDa subunits that assemble randomly into active tetramers (5). The expression of the different genes varies from one tissue to another and within a tissue during development (6). One subunit type (B) occurs predominantly in tissues such as heart and liver where lactate is metabolized. In contrast to the predominant heart isozyme (LDH-B4), the predominant skeletal muscle isozyme (LDH-A4) is catalytically adapted for tissues like anaerobic skeletal muscle or red blood cells where lactate is produced (7). Testes contain a third type (LDH-C4). All three are the result of successive duplications of an ancestral gene (8, 9) and thus are paralogous. Where multiple subunits are present, hybrid isozymes (e.g., A3B, A2B2, and AB3) occur. The distinctive catalytic properties and tissue-specific patterns of subunit production suggest functional differentiation.
Recognizing that hypotheses of function and evolutionary relationships could be tested by detailed comparative studies, Nathan O. Kaplan, then chairman of the Biochemistry Department at Brandeis University, embarked on a major program on the comparative biochemistry of LDH (3, 10, 11). Typically he would put first-year graduate students under the supervision of a postdoctoral fellow and give them the task of purifying and characterizing LDH from the heart or skeletal muscle of some exotic animal obtained after death from a zoo. Notable among the postdoctoral fellows attracted to Kaplan's laboratory was Allan C. Wilson (see Case Study 3), who used immunological cross reactivity as a measure of structural similarity in LDH.
For initial group discussion: Make a list of the terms and concepts in the preceding introduction that you don=t understand or feel you need to learn more about. Use the textbooks and other resources available in the classroom to help clarify your understanding. Each group needs to turn in a list of the terms that were discussed before proceeding to the assignments for future class periods.
References
1. Darwin, C. (1859) The
Origin of Species.
2. Markert, C. L. (1968)
"The molecular basis for isozymes" Ann. N.Y. Acad. Sci. 151,
14-40.
3. Kaplan, N. O. (1964)
"Lactate dehydrogenase--Structure and function" Brookhaven Symposia
in Biology 17, 131-153.
4. Eventoff, W., Rossmann,
M. G., Taylor, S. S., Torff, H-J., Meyer, H., Keil, W. and Kiltz, H-H.
(1977) "Structural
adaptations of lactate dehydrogenase isozymes" Proc. Natl. Acad.
Sci. USA 74, 2677-2681.
5. Markert, C. L., Shaklee,
J. B. and Whitt, G. S. (1975) "Evolution
of a gene" Science 189, 102-114.
6. Cahn, R. D., Kaplan,
N. O., Levine, L. and Zwilling, E. (1962) "Number
and development of lactate dehydrogenases" Science 136,
962-969.
7. Flores, R. (1979) "Mode
of action of lactate dehydrogenase isozymes" TIBS 4(2), N32-N33.
8. Li, S. S-L., Fitch, W.
M., Pan, Y-C. E. and Sharief, F. S. (1983) "Evolutionary
relationships of vertebrate lactate dehydrogenase isozymes A4
(muscle), B4 (heart), and C4 (testes)" J.
Biol. Chem. 258, 7029-7032.
9. Crawford, D. L., Constantino,
H. R. and Powers, D. A. (1989) "Lactate
dehydrogenase-B cDNA from the teleost Fundulus heteroclitus: Evolutionary
implications" Mol. Biol. Evol. 6, 369-383.
10. Kaplan, N. O. (1986)
"Experiences in biochemistry" in Comprehensive Biochemistry, Vol.
36, pp. 255-296 (M. Florkin and E. H. Stotz, eds.) Elsevier Science Publishers,
Amsterdam. See also Protein Sci. 4, 311-325 (1995), especially
pages 323-324.
11. Kaplan, N. O. (1965)
"Evolution of dehydrogenases" in Evolving Genes and Proteins (V.
Bryson and H. J. Vogel, eds.) pp. 243-277, Academic Press, N.Y.
12. Wilson, A. C., Cahn,
R. D. and Kaplan, N. O. (1963) "Functions of the two forms of lactate dehydrogenase
in the breast muscle of birds" Nature 197, 331-334.